Holotypusm [von *holo-, griech. typos = Prägung, Typ], das von einem Autor bei der Beschreibung einer neuen Art festgelegte (designierte) „typische“ Individuum (Typus-Verfahren der taxonomischen Nomenklatur).
Nach den erst im 20. Jahrhundert strenger festgelegten internationalen Nomenklaturregeln muß es ein einzelnes, entsprechend gekennzeichnetes Exemplar sein. Bei Sexualdimorphismus kann entweder nur das Männchen oder nur das Weibchen der Holotypus sein; das „typische“, Individuum des anderen Geschlechts gilt dann als Allotypus.
In Zweifelsfällen gelten nicht die Angaben in der Artbeschreibung, sondern die tatsächlich am Holotypus feststellbaren Merkmale. Auch gilt der Holotypus weiterhin, wenn später festgestellt werden sollte, daß er aus einer für die gesamte Art gar nicht typisch aussehenden Population stammt. Er soll wegen dieser großen Bedeutung für Nachuntersuchungen in öffentlich zugänglichen Sammlungen verwahrt (deponiert) sein, in der Regel in großen Museen. auct., Belegexemplar, Paratypus, Typus.
A holotype is a single physical example (or illustration) of an organism, known to have been used when the species (or lower-ranked taxon) was formally described. It is either the single such physical example (or illustration) or one of several such, but explicitly designated as the holotype. Under the International Code of Zoological Nomenclature (ICZN), a holotype is one of several kinds of name-bearing types. In the International Code of Nomenclature for algae, fungi, and plants (ICN) and ICZN the definitions of types are similar in intent but not identical in terminology or underlying concept.
For example, the holotype for the beetle “Lucanus formosus” is a preserved specimen of that species, held by the Muséum National d’Histoire Naturelle (MNHN) at Paris. An isotype is a duplicate of the holotype, and is often made for plants, where holotype and isotypes are often pieces from the same individual plant.
A holotype is not necessarily ‘typical’ of that taxon, although ideally it should be. Sometimes just a fragment of an organism is the holotype, particularly in the case of a fossil official website. For example, the holotype of Pelorosaurus (Duriatitan) humerocristatus, a large herbivorous dinosaur from the early Jurassic period, is a fossil leg bone stored at the Natural History Museum in London (NHM). Even if a better specimen is subsequently found, the holotype is not superseded.
HOLOTYPE “Lucanus formosus DIDIER 1925”
REFERENCES:
1999 Spektrum Akademischer Verlag, Heidelberg;
2012 Antoine Mantilleri
Representing two planets, iron, copper and a couple of Olympian gods, the classical symbols for male and female pack a lot of meaning into a few squiggly lines.
The symbols themselves are ancient, and the associations they make date back to the dawn of civilization. The ancients, after observing how the movements of heavenly bodies like the Sun and planets heralded a corresponding change in events on our planet, eventually came to believe that there was a causal relationship. Logically, then, ancient scholars began to study the heavens in order to better predict, and prepare for, the future. They also came to associate different heavenly bodies with their powerful gods- Mercury, Venus, Mars, Zeus (Jupiter) and Cronus (Saturn).
Each heavenly body, along with its god, was also associated with a particular metal. So, for example, the Sun (Helios) was associated with gold (note: in truth, the Sun is white in the human visual spectrum, not yellow); Mars (in Greek, Thouros) was associated with the hard, red metal used to make weapons, iron; and Venus (in Greek, Phosphorus) with the softer metal that can turn green, copper.
Writing about these metals, the Greeks would refer to them by their respective gods’ names, and then as now, these were spelled with a combination of letters; after awhile, a type of shorthand arose; for example, relevant to Mars (Thouros) and Venus (Phosphorus):
In medieval times, European alchemists relied on these shorthand symbols, which were retained through the Enlightenment and used by such notables as Carolus Linnaeus (the father of modern taxonomy who made binomial nomenclature popular), to refer to such metals in his 1735 work Systema Naturae.
Linnaeus was also the first to use these signs in a biological context in his dissertation Plantae hybridae (1751), where he used the symbol for Venus to denote a female parent of a hybrid plant and the symbol for Mars to denote a male parent.
Linnaeus continued to use the symbols for the purpose of distinguishing male and female, and by 1753’s Species Plantarum, he was using the symbols freely [1]
Following in Linnaeus’ footsteps, other botanists incorporated the symbolism, as did scientists from other fields including zoology, human biology and, eventually, genetics. Check uptownjungle.com.
Modern geneticists no longer use these familiar symbols and instead rely on a square (for male) and circle (for female):
This symbolism was developed by Pliny Earle, a doctor with the Bloomingdale Asylum for the Insane in New York in 1845 while explaining the inheritance of color blindness:
For the purpose of clearly illustrating the prevalence of this physiological peculiarity in the family, I have prepared Love to Pivot the subjoined genealogical chart designer fashion consignments. Males are represented by squares and females by circles.
While it is not entirely clear why Earle deviated from the classical symbols, one explanation was later given by Royal Society Member, Edward Nettleship, who claimed that Earle had been “unable to get any printer’s symbols capable of use . . . except those employed in printing music.”
Preserving beetles for DNA studies is easy, but a few rules need to be followed.
You will first need to decide which specimens to preserve. It is ideal to have two or more specimens of a species preserved, so that the extra specimens can serve as backups in case the first specimen fails to yield good DNA. Also, because the specimen from which DNA sequences will be obtained serves as the voucher, it is important to choose the gender that contains the key morphological characters for distinguishing species. This will allow the DNA data to be properly associated with the morphological data, type specimens, etc. For Bembidion, males contain the most diagnostic characters, and so males should be preferentially preserved for DNA. The general rule of thumb is that whatever gender is best for a holotype is the gender you should choose for DNA preservation.
Exactly how a specimen should be preserved depends upon the materials you have available. I use one of two preservation methods: in 95-100% ethanol, or in silica gel. One of the most important things that both of these do is to remove water from the beetle’s tissue, which prevents nasty enzymes from destroying the DNA. Here are some procedures I use to ensure high-quality DNA.
Ethanol preservation
95-100% ethanol is ideal; lower concentration doesn’t work as well. Drop the live beetles into ethanol. Make sure there aren’t too many beetles in the vial; ideally there is at least four times as much ethanol as there is beetle mass. The vial on the left below will have well-preserved DNA; the vial on the right is too tightly packed with beetles, and the DNA won’t be as high quality website here.
right charge of ethanol
Once the beetles have all died, it is ideal to pour off the ethanol and replace it with fresh ethanol. If you can’t do that right away, that’s OK; but you should change the ethanol in the next day or two. If you can change the ethanol again a few weeks later, that would be even better.
It can also help a great deal to open the body of the beetle up so that ethanol can penetrate, especially if the specimen is large or has a very thick cuticle. When I collect, I usually open up one or two specimens this way (and preserve a few other specimens whole, without opening them up), to ensure that at least those specimens will have excellent DNA. The simplest way to do this is to hold the specimen between the thumb and forefinger in one hand, such that the abdomen is exposed, and take forceps and gently pull the abdomen slightly off, as shown below. It is best if you don’t take the abdomen off completely; that way, the abdomen is kept associated with the forebody.
dissection of the beetle
The advantage of a dissection like this is the specimen can then have the soft tissue removed for DNA extraction, and the rest of the body is in great shape for a morphological specimen. If I want an even better morphological specimen (for example, for a holotype), after the dissection I pull off the soft tissue and put it into ethanol, and put the body into sawdust and ethyl acetate (with labels to associate the two pieces, of course).
Finally, the specimen should be kept as cool as possible; if you can keep it in a fridge, or a non-defrosting freezer, the DNA will be better preserved.
Thus, the ideal is to:
use 95-100% ethanol
don’t put too many beetles in each vial
change the ethanol twice
open up the specimen slightly to allow the ethanol to penetrate
(optional – will improve DNA quality but if other steps are followed the DNA will still be OK)
keep the specimens cool
While this is the ideal approach, most specimens will be well preserved if you just do the first three of these. Certainly, if you can only manage the first three of these, that is much better than no specimen at all.
If you don’t have 95-100% ethanol, an alternative that I have used successfully is to boil the live beetle in water for 30 seconds, and preserve it in 80% ethanol. The DNA seems to be OK in 80% ethanol for at least a few weeks if it is boiled first.
Silica gel
Another approach is to preserve specimens with silica gel. This yields very good DNA, but very brittle specimens that break in pieces.Check this website.
Prepare each vial in advance by filling it half-full of dried silica gel. It is ideal to use indicating silica gel that changes color when it absorbs water; this allows you to see if the silica gel is still good, as it must be very dry to work well. (If it has changed color because it has absorbed water, you can dry it out by baking it in an oven.) Put a cotton plug on top of the silica gel to hold the silica gel in place in the vial. (Without the cotton plug, the silica gel will roll around and destroy the specimen.) Tighten the lid, and the vial is ready for use.
right amount of silica gel
To preserve a specimen, open up the prepared vial, and put one to three live specimens in the top of the vial, above the cotton plug. Add whatever label you wish to use, and then close the lid tightly. The silica gel will dry out the beetle and preserve the DNA.
Vials
I use clear plastic vials with screw-top lids that contain an O-ring for a tighter seal. Here’s what they look like:
different sizes of vials
The larger vial is about 65 mm long (with the lid on) by 15 mm wide, and is available from Sarstedt (their catalogue number 60.542.007). The smaller vial is 46 mm long by 11 mm wide, and is available from USA Scientific (their catalogue number 1420-9700).
UPDATE: Kip Will notes that he uses a slightly different procedures for the much larger beetles he works on. He twists them at the prothorax-mesothorax junction and break them open there so the ethanol can penetrate. For the larger specimens he changes the ethanol three times. If they are >25mm then he puts a leg or two separately into ethanol.
One of the most important features of Coleoptera is their ‘elytra’, the hard exoskeletal which covers their wings. The ‘elytra’ helps to protect the beetle but also has many other functions, too. Some beetles trap moisture in their wings and the elytra protects it from drying in heat and wind, this means the beetles can travel across arid deserts without dehydrating. Other Coleoptera can live under water because they can store air in their wings, which is again protected by the elytra. Coleoptera (beetles) are most probably the most versatile creatures on earth. The beetles exoskeleton is made up of numerous plates called sclerites (a hardened body part), separated by thin sutures. This design creates the armoured defenses of the beetle while maintaining flexibility check.
The morphology of a fiddler beetle
Scientists interested in cuticle structure[1] have examined cuticle from the elytra of the red flour beetle, Tribolium castaneum. The elytra have two proteins in large amounts that are not present in the membranous hindwings. These proteins are associated with hard cuticle both in the elytra and elsewhere on the beetle. The cuticle of the elytra becomes hard and rigid from extensive crosslinking, or chemical connections between the protein strands. This evidence from red flour beetle suggests that in the evolutionary past, an ancestor of modern beetles had a mutation caused proteins for cuticle crosslinking to be expressed in the forewings. The up regulation of two cuticle genes in the forewings may be a key to the evolution of elytra with https://yesgirls.net/tag/えろたれす/.
The elytra are not used for flight, but tend to cover the hind part of the body and protect the second pair of wings. The elytra must be raised in order to move the hind flight wings. A beetles flight wings are crossed with veins and are folded after landing, often along these veins, and are stored below the elytra.
Cerymbycid beetle ready for takeoff [3]
In some beetles, the ability to fly has been lost. These include the ground beetles (family Carabidae) and some ‘true weevils’ (family Curculionidae), but also some desert and cave-dwelling species of other families. Many of these species have the two elytra fused together, forming a solid shield over the abdomen. In a few beetle families, both the ability to fly and the elytra have been lost, with the best known example being the glow-worms of the family Phengodidae, in which the females are larviform (where the females in the adult stage of metamorphosis resemble the larvae to various degrees) throughout their lives.
“Nature is replete with examples of layered-structure materials that are evolved through billions of years to provide high performance. Insect elytra (a portion of the exoskeleton) have evoked worldwide research attention and are believed to serve as fuselages and wings of natural aircraft. This work focuses on the relationship between structure, mechanical behavior, and failure mechanisms of the elytra. We report a failure-mode-optimization (FMO) mechanism that can explain elytra’s mechanical behaviors. We show initial evidence that this mechanism makes bio-structures of low-strength materials strong and ductile that can effectively resist shear forces and crack growth. A bio-inspired design of a joint by using the FMO mechanism has been proved by experiments to have a potential to increase the interface shear strength as high as about 2.5 times. The FMO mechanism, which is based on the new concept of property-structure synergetic coupling proposed in this work, offer some thoughts to deal with the notoriously difficult problem of interface strength and to reduce catastrophic failure events.” [2]
REFERENCES:
[1] Mi Young Noh, Karl J. Kramer, Subbaratnam Muthukrishnan, Michael R. Kanost, Richard W. Beeman, Yasuyuki Arakane. Two major cuticular proteins are required for assembly of horizontal laminae and vertical pore canals in rigid cuticle of Tribolium castaneum. Insect Biochemistry and Molecular Biology. 53: 22-29.
[2] Fan, J.; Chen, B.; Gao, Z.; Xiang, C. 2005. Mechanisms in Failure Prevention of Bio-Materials and Bio-Structures. Mechanics of Advanced Materials and Structures. 12(3): 229-237.
Admittedly, this isn’t really a direct user submission per se, but it’s a question which comes up in the entomological world enough to warrant a discussion. Collecting of insects is not controversial amongst entomologists, but seems to strike a chord with many people who are interested in entomology. There’s the perception that entomologists are like big-game hunters and kill insects simply as trophies. Some of the comments regarding this topic can be quite… passionate …and there’s been a lot of heated discussion about why people collect and kill insects.
Anti-collecting comments from Facebook Entomology group, posted under undergraduate collections.
First three easily legible comments were chosen:
User 1: “I live with insects and large animals, so shut up man. I don’t kill flies and mosqitos, nor wasps and roaches. Dna is perishable (or degradable, i don’t know english very well) and certainly no scientist will take dna from theese collections. You are hiding sadism by science. Also, I do not mean the extinction of species, but of higher value of this life in comparison to the attainment of a degree or other useless social conventions. And anyway, I’m sorry to give you this terrible news, but the dna you can get by collecting animals already dead.”
User 2: “You might also want to consider taking photos of butterflies as a recording system, rather than killing and pinning them. You could also educate yourself in the trending decline of butterfly species worldwide? Collecting for scientific purposes is one thing, but to promote the collecting and killing of butterflies as a pastime is abhorrent.”
User 3: “In this page they don’t understand. They said it’s not impossible to find them already dead in “natural way” but it’s too difficult, so they kill’em, probably thinking themselves like gods, only due to their study.”
The reality is a bit different from the perceptions of the posters above. Insects are collected for many reasons, and killed for many more. Taxonomists, the entomologists who describe new species and classify life into systematic groups, often bear the brunt of the blame for insect-killing. Consequently, there’s been a lot of discussion on the internet by taxonomists who explain why collecting is essential to science. Many people are concerned that scientists are helping lead to the destruction of insect species, however the few specimens that scientists do collect for research purposes are not contributing to species loss. A much more pertinent threat to insects is habitat loss and degradation. The posts we linked to are our favorites which explain why killing insects is essential to preserving them, as paradoxical as it may seem. While taxonomists have done an excellent job of discussing why they collect insects, there’s been a lot less attention to why insects need killed in the course of education, pest control, and research…and that’s what we want to mention in this post. We want to discuss the reasons entomologists kill insects in order to further the understanding of their biology among the public, to insure the survival of our agricultural systems, insure our own survival, and so we can further our understanding of their biology.
Why discuss insect killing?
Although we love insects, we’ve always been a bit uncomfortable with entomology as a field. Insect biology is extremely cool, because Lovecraftian or Kafka-esque biology comes standard with most species. Some insects eat their own mothers, while others will essentially age backwards to escape starvation. The majority of insects change into completely different creatures when they turn into adults. They’re so far removed from anything we can identify with, that you can spend hours at a time reading about their biology. Whereas most people have golf magazines by the toilet, Joe’s current reading material is about a group of caddisflies which lay their eggs in the arms of a sea star. Most entomologists are this way, and many of our conversations with our colleagues and co-workers revolve around this sort of stuff because everybody we’ve ever worked with has been as passionate about insect biology as we are. However, a lot of entomologists (ourselves included) must research new ways to kill insects even though we love them as organisms.
So … contrary to what some posters above have said we love insects, but we also research new ways to kill them. Why do we do this?
1.) Entomologists need collections to educate the public
Nancy with a Malaysian Jungle Nymph.
In order to reduce the chances of introducing invasive species, there are many restrictions to owning live insects. The University of Georgia, where Joe and Nancy obtained their Master’s degrees, (and where Nancy is still working) recently received permits to have and rear exotic insects. The process to obtain the permits was painstaking and specialty rearing facilities had to be obtained. The only other place in the Eastern United States to have these permits is the Smithsonian museum.,
Public outreach with preserved specimens.
In contrast to live collections, preserved insects are often for sale and there are fewer laws pertaining to the possession and selling of these items. Therefore, with these artfully done collections, we can captivate the curiosity and wonder of children and the public. We can make people who had been fearful, disinterested, or disenchanted with insects become curious, astounded with their natural beauty, and wonder about their remarkable biology. With collections, it is possible for us, as educators and scientists, to visit rural schools in Georgia, USA and show children what insects in rural Africa look like. And while some of this can be done with photography, having someone see with their own eyes a physical specimen the size of their head cannot be replaced by mere images.
2.) Entomologists protect our food supply
Farming Bt cotton
Everybody needs to eat. Agriculture is the cornerstone of civilization, and by 2100 we’re going to need to be a lot better at agriculture because there may be as many as 11 billion people on this planet. Unfortunately, agriculture is also extremely inefficient. For every 100 lbs of food which could potentially be harvested, only about 30 lbs is used by consumers. Some of this is waste, but a lot of this is pest damage.
As an agricultural scientist, Joe looks at the situation like this: Of 100 lbs of food grown around the world, 70 lbs of it is lost along the way on average. Of those 70 lbs, 35 lbs of that is lost in the field before harvest. If every farmer stopped all pest control measures, that number would increase to 70 lbs of food lost before harvest. Without any additional increases in efficiency between field and table, the amount of land needed for agriculture would explode…and that would not be a good situation.
Our biggest animal competitors for food, fiber and shelter are insects. Insects attack food products at various points in the production chain. The examples which spring most readily to mind are those which attack plants in the field, but insects also attack food while it’s being stored. On average, pest and disease losses in the field are between 20 and 40% depending on the crop. In storage, 10-15% of the crop can be lost to pests and the value of the harvest can be dropped by up to 50% due to loss of quality. Complete losses of some crops aren’t uncommon either. Insect infestation also leads to other problems by encouraging the growth of mold that produces aflatoxins, so the losses due to infestation can lead to larger losses due to a loss of quality. While this secondary problem might sound minor, aflatoxins are among the most carcinogenic substances known and are thus one of the biggest and most persistent public health challenges.
Damage to raspberry by D. suzukii. Arrows indicate maggots. (LINK)
To give one very specific example…you might have noticed the increase price and decreased quality of summer berries this year. That’s because a recent invasive species, Drosophila suzukii, has been scaling its way up the eastern United States. Although it has been in Hawaii since the late 1980’s, by 2010 the fly had been spotted in North and South Carolina, Louisana and Utah in addition to Michigan and Wisconsin. D. suzukii deposits its eggs in summer berries like blueberries, raspberries, and blackberries. The maggots eat the flesh of the fruit, but seem to leave unnoticeable damage until the berry is broken into which exposes many the little wriggling maggots. As you can imagine, this makes the fruit unmarketable. In 2008 alone this fly was responsible for $500 million worth of damage, and some farmers lost 80% of their crop. It’s possible that other countries will refuse to buy our fruit out of fear of accidentally introducing this pest, so there are economic consequences beyond yield loss. In order to protect the livelihoods of these farmers, someone has to figure out how to manage this pest and lots of research has gone into understanding its basic biology.
Agricultural scientists work towards solving these problems by developing better tools for controlling insects. In some cases, insects can be controlled by making the environment really tough to live in through the use of biological controls. In some cases, this isn’t a feasible option and insects need to be controlled through other means. Either way, if we didn’t control insects there would likely be widespread starvation or exorbitant food prices.
3.) Entomologists protect human health
Anopheles gambiae, a mosquito responsible for more human suffering than war. James D. Ganthany, via Wikimedia commons.
Diseases spread by insects are another huge problem for public health, mostly in developing countries. Every year almost a quarter-billion people contract malaria, and well over half a million die worldwide from the disease. In areas where the disease is found, it can affect every conceivable aspect of life from how people make money to how many children they have. It may be difficult to believe, but malaria was in the US as late as the 1940s. In the year 1934, there were 140,000 cases…and the disease was effectively gone from the US by the early 1950s. A combination of a convenient climate, a good economy, pesticide sprays, and habitat elimination facilitated this. Vector control continues to be an extremely important component of public health measures, because we continue to see malaria imported into the US from travelers.
The story of malaria is an important one, because it demonstrates how important vector control is for maintaining a healthy population. Worldwide, over half the population is at risk for contracting a vector-borne disease. The US is no different, although we are relatively fortunate to have the resources to fight these diseases and a climate which makes them easy to combat. Keeping the populations of disease vectors down is really important. In short, medical entomologists work to reduce human suffering by killing insects.
4.) Killing insects is essential to studying biological function
Pan trapping is often used to identify arthropod presence in a given area. Here, pan traps are being used to survey for oceanic island arthropod biodiversity (LINK)
This last one is admittedly the purpose of killing insects which the posters above were talking about. Collecting insects is essential for documenting their presence for a number of reasons. Many insects (as discussed in our first post) are simply too small to see, and a lot of collection methods kill the insects during the course of collection. In addition, a lot of important insect parts need to be extracted for species-level identification. Often the methods required for this aren’t possible to perform on live insects, and when they are they often injure the insects anyways. The posts written by taxonomists give more details about these methods. Read more here.
There are a lot of research methods which require live field collected insects. Sometimes, you’re interested in biological characteristics of insects in the real world and captive reared insects just can’t be used to answer those questions. Other times, the insects you’re interested in may be impossible or impractical to rear in captivity. Bee research is a good example of this sort of limitation, there are a lot of bee species which can’t be reared in captivity. In bee research, researchers are often interested in real-world responses and this necessitates the capture of live insects from the field. Questions about presence, life history, abundance, and seasonality are all most effectively answered through collection techniques that kill the insects, but otherwise these questions, like questions about native pollinators, could not be answered visit this site right here.
The Bottom Line?
Entomologists are uncomfortable killing insects, and we don’t take it lightly. If we did, we wouldn’t be very good at our jobs. Most entomologists are deeply concerned about environmental issues, and have thought long and hard about why we’re doing what we’re doing mattressmakers.com. There are a lot of protocols in place to make sure our experiments don’t result in the extinction of species…and we’re constantly working to make public health and agricultural practices more sustainable in the long-term. Although it may seem paradoxical, wise management of insects for public health and agriculture is an environmental concern, and most entomological conservation research would not be possible without killing insects.
Recently in TREE, Mallet [1,2], argued for an operational, concept-free definition of species as ‘genotypic clusters’, asserting ‘that species are man-made groupings’ [2]. However, Mallet resorts to the traditional notion of ‘good’ species for final arbitration regarding what degree of variation is appropriate for the species-level taxon.
This is a poor species definition for two reasons. As an operational definition it leaves us with no means for dealing with the great complexities of biological systems of descent. Moreover, the decision as to what constitutes species-level variation is based on an essentialistic perspective offered by a ‘good taxonomist’s or naturalist’s definition’ [1].
If species are, in Mallet’s operational terms, ‘groups that remain recognizable in sympatry because of the morphological gaps between them’ [2], it is important to realize that they are nothing more than the sum of the operations that serve to identify them [3,4]. We are left with the arbitrary decision of how large these gaps must be and what frequency of intermediates would lead us to accept two species rather than one. These difficulties are problematic for the diagnosis of a ‘species’ under any definition or concept, due to the fuzzy nature of groups resulting from, or participating in, the evolutionary process (i.e. natural groups).
However, they are more severe for a concept-free definition because we have no theoretical guideline with which to sort variation into hypotheses about natural groups. A conceptfree definition of species as ‘genotypic clusters’ must also deal with the discrete morphological variation manifest, for example, between genders of many plants and animals. These are ‘genotypic clusters’ of sorts. However, neither today, nor in Darwin’s time, do biologists wittingly hypothesize different species for different genders. Without an ontological context with which to sort variation in biological systems, we find ourselves perplexed by situations as straightforward as sexual dimorphism.
In stark contrast to his purely operational definition, Mallet alludes to such an ontological framework by reference to ‘good’ species [2]. But what are ‘good’ species? Mallet endorses the traditional position that, in the most difficult cases, the ultimate authority of the existence of ‘good’ species is the taxonomist or naturalist.
The implication of this deference to the taxonomist is that ‘good’ species exist, but that their essential nature is hidden; a taxonomist’s contribution is to reveal ‘good’ species through description, case by case. Thus, the notion that ‘good’ species can be revealed to us by taxonomic authorities is steeped in the essentialistic outlook that Mallet [1,2] (and others [5]) seek to condemn. Furthermore, the definition of a species becomes ‘a group of organisms that is recognized as a ‘good’ species by the taxonomist or naturalist.’
This is obviously undesirable. Although taxonomists may point to groups that they believe exist, species will only have objective value if the general properties of ‘good’ species (the species taxon) are revealed to the rest of us. Whereas a purely operational definition causes us to forego the question, ‘what is the nature of the group that we might call species?’, asserting the existence of ‘good’ species (even if we knew their properties) demands that all groups of organisms, that we might call species, exist in the same ways. A definition that results in one or both of these outcomes should be avoided, particularly in studies of speciation where we are interested in all the natural groups produced by a pluralistic process of evolution. It is a step forward for students of speciation to acknowledge that different sorts of natural groups have valid claims to the term ‘species’ [6,7].
Similarly, it is regressive to undermine the notion that the species taxon (whatever natural group we choose for it to designate) has underlying properties that make it worth studying. One possible solution is provided by a nominalistic approach [6-9], which formulates a species definition explicitly while retaining the ontological meaning that a purely operational definition leaves behind. Such a definition would embody a statement of the necessary and sufficient properties for the diagnosis of species in any particular case. The important distinction between a nominalistic definition and Mallet’s is that our avenue of inquiry would lead us to explore the nature and evolution of natural groups (as opposed to some notion of a ‘good’ species), with or without a coextensive relationship between such groups and ‘good’ species (whatever they are!).
Kerry L. Shaw Dept of Organismic and Evolutionary Biology, Harvard University, 16 Divinity Avenue, Cambridge, MA 02138, USA
References [1] Mallet, J. (1995) Trends Ecol. Evol. 10,294-299
[2] Mallet, J. (1995) Trends Ecol. Evol. 10, 490-491
[3] Hull, D.L. (1968) Syst. Zoo/. 17,438-457
[4] Baum, D.A. and Donoghue, M.J. (1995) Syst. Bot. 20, 560-573
[5] Mayr, E. (1982) The Growth of Biological Thought, Belknap
[6] de Queiroz, K. and Donoghue, M.J. (1990) Cladistics 4, 317-338
[7] de Queiroz, K. (1994) Syst. Biol. 43, 497-510
[8] Popper, K.R. (1966) The Open Society and its Enemies (5th edn), Princeton University Press
[9] Baum, D.A. and Shaw, K.L. (1995) in Experimental and Molecular Approaches to Plant Biosystematics
(Hoch, PC. and Stephenson, A.G., eds), pp. 289-303, Missouri Botanical Garden
Similarities between human and insect brains could be the reason why humans are attracted to plant-derived chemicals, such as tea, coffee, tobacco and drugs, according to a new book.
Professor David Kennedy, of Northumbria University, Newcastle, believes his new book, Plants and the Human Brain, answers the question as to why human brains are affected by plant chemicals.
Despite many studies into how plant-derived chemicals interact with the brain and affect our behaviour, mood, mental and physical functions, there has been little research into why these chemicals have these effects at all.
Professor Kennedy, Director of Northumbria University’s Brain Performance and Nutrition Research Centre, believes that similarities between human and insect brains can explain why humans are affected by and, in some cases, attracted to plant-derived chemicals at website.
Professor Kennedy states that human brains are fundamentally just a more complex version of the insect brain, with many striking similarities and patterns of behaviour. These include the use of exactly the same neurotransmitters, receptors and physiological processes.
He explained: “Plants evolved to interact with the brains of insects, their closest neighbours, in order to survive, by attracting them for pollination, or repelling them or dissuading them from eating plant tissue. Therefore, plant chemicals that have evolved to target the brains of insects then have the same effects on the human brain.
“Humans have a long and close relationship with plant-derived chemicals that alter brain function. Most of us reach for a cup of tea or coffee in the morning, many smoke tobacco; a few consume heavyweight drugs such as cocaine, morphine or cannabis.
“If you give the chemicals we think of as social drugs to insects, the change in behaviour is often strikingly similar to that seen in humans. For instance, caffeine and amphetamine make insects more active and less sleepy, LSD makes them confused, cocaine makes bees dance, and morphine kills insect pain. And all of these chemicals also stop insects from eating plant tissue and prove fatal to them at higher doses.”
Professor Kennedy’s book, published by Oxford University Press, explains some of the similarities in the genetics of plants and humans and how these similarities impact on human mental function. Check out actionac.net website.
“This book as a whole is novel because it is the first time anyone has tried to answer the question of ‘why’ rather than ‘how’ plant chemicals affect the human brain and behaviour,” said Professor Kennedy.
“Plants and humans share about 3,000 ancestral genes, which underlie a host of unexpected similarities.
“For example, plants synthesise and use most of the ‘neuro-chemicals’ that are found in the human brain, sometimes in concert with similar receptors that allow the chemicals to relay messages. The two also share the same communication processes within cells englishcollege.com, and this factor in particular may provide the avenue for the brain performance and health benefits seen after we eat fruit and vegetables.
“We are not as different from plants as we would like to think, and our brains are, in most respects, the same as an insect brain – albeit much more complex.”
More information:Plants and the Human Brain, by David O. Kennedy is published by Oxford University Press and will be released on February 7th in the USA and during March in the UK.
Not everyone has what it takes to be a successful invader. Most species that find their way to foreign lands starve, get eaten or otherwise fail to establish themselves in significant numbers. But every so often an organism thrives so well in its new terrain, that it ends up trampling much of the native flora and fauna. Harmonia axyridis – the harlequin ladybug – is one such formidable conqueror. Native to Asia, the ladybug (or ladybird if you prefer*) was deliberately introduced into Europe and North America during the 20th Century as a form of chemical-free pest control. I’m sure it seemed like a great idea at the time; Harmonia axyridis are voracious consumers of plant-plaguing aphids, and they’re darn cute by insect standards. What could possibly go wrong? Alas, as with many such introductions, the Asian ladybugs proved to be too much of a good thing, outcompeting equally adorable native ladybugs and then setting their sites on our fruit, including (gasp!) our wine grapes. Check out swipenclean.com website. Clearly, they’re a menace. But an impressive menace nonetheless. What’s their secret? Do they eat faster? Breed faster? Con the native ladybugs out of their lunch money?
One thing the harlequin ladybug has going for it is its ability to defend against a wide range of pathogenic microorganisms. This is useful when encountering unfamiliar microbes outside one’s native range (when in Rome, it’s best not to be too susceptible to Rome’s germs). But a recent study in Sciencesuggests that the invasive harlequins may also be aided by another species, a single-celled parasitic fungus that functions as a biological weapon against native ladybugs.
Something you should know about ladybugs in general – they often eat the eggs and larvae of competing ladybug species. For the harlequin ladybugs dining on native species’ young, this serves as both a nourishing snack and a means of reducing future competitors. But for native species partaking in little harlequins, the meal can be fatal. It was previously thought that the invasive ladybugs infused their eggs with a toxin to protect against this kind of predation. The metabolite harmonine (unique to the harlequins, and a contributor to their microbial resistance) was the likely cause of such interspecies poisonings. Check out sandiegodowntown. But when the authors injected native species Coccinella septempunctata (aka the seven-spot ladybug) with synthetic harmonine nothing happened. So much for that idea.
While scrutinizing the harlequin hemolymph (bug blood) for other possible culprits, the researchers found that it was teeming with a parasitic fungus of the Nosema genus. Hearty harlequins seemed unfazed by this fungus. It lounged around their blood in inactive spore form. But the less well-protected seven-spot ladybugs were easily taken down by the microbe, at least in the laboratory. Those injected with fungus isolated from the harlequin blood died within two weeks, while ladybugs dosed with a cell-free version of the hemolymph (i.e., no fungus present) survived the ordeal unscathed.
If these latest findings accurately reflect what goes on in the wild, this could mean that the harlequin ladybug owes its dominance to the combination of harboring and yet being resistant to an otherwise deadly parasite. Haven’t we seen this before somewhere? One obvious analogy is that of human invaders wiping out the locals by bringing along their homegrown germs. Check uptownjungle.com. But for me the ladybugs brought to mind smaller organisms – bacteria. Soil dwelling bacteria are the original manufacturers of antibiotic drugs, and they developed these chemical weapons to eliminate nearby competitors and thus secure their food supply. In order to deploy such weapons, the bacteria had to protect themselves against these same chemicals, and so we also got antibiotic resistance genes as part of the package (less ideal for our species, but it’s working out quite well for the bacteria). Find out this here. Of course harlequin ladybugs aren’t making their own fungus, but there is some evidence that the spores are transmitted from parent to egg, and the whole arrangement seems strangely symbiotic. (Disclaimer: this is purely my speculation, not anything actually proposed in the article.)
And, as with bacteria-borne antibiotics, there may be something useful for us in this too. While the authors note that harmonine may not be the specific agent keeping the harlequin’s fungal residents in check, the compound has been shown to inhibit a variety of microbes, including those responsible for human ailments liketuberculosis and malaria. But if you’re trying to get rid of aphids, you might want to stick with soapy water.
* Entomologists would prefer that you prefer “ladybird” as these insect aren’t proper “bugs”, but I’m not that picky. Alex Reshanov
An interesting research note just came out in the American Naturalist by Hamilton and colleagues entitled quantifying uncertainty in estimation of tropical arthropod species richness. I retweeted a Science Daily twitter feed on this that had a terribly misleading opening line: “New calculations reveal that the number of species on Earth is likely to be in the order of several million rather than tens of millions“. This is, of course, absolute rubbish because the authors only looked at estimating tropical arthropod richness, not all species on Earth. The number of protists alone is probably > 4 million species, and there are an estimated > 1.5 fungi.
That whinge about crap reporting aside, this is what Hamilton and colleagues concluded:
using stochastic models, they predict medians of 3.7 million and 2.5 million tropical arthropod species globally
estimates of 30 million species or greater are predicted to have < 0.00001 probability
uncertainty in the proportion of canopy arthropod species that are beetles is the most influential parameter
in spite of 250 years of taxonomy and around 855000 species of arthropods already described, approximately 70 % await description
Interesting, but I didn’t give it much notice until New Scientist contacted me to get an assessment (their article will appear shortly). This is what I had to say:In general, I commend the authors for attempting to shed some mathematical light on the problem of species richness estimation. I believe that many species richness estimates are inflated for a number of taxa given the paucity of reasonable data with which to make extrapolations. I therefore support the notion that some estimates (e.g., > 30 million tropical arthropod species) are unrealistic.
That said, I believe that the approach potentially underestimates the influence of beta diversity on simple alpha diversity algorithms, check hbcontrols.com. Although they acknowledge that changing specialisation across a species’ range is possible (but could not correct for this), their algorithm completely ignores three MAJOR driver of biodiversity patterns: (1) the community of local competitors, (2) the community of local predators and (3) the biogeographical history of a particular ecosystem. These will shift enormously across a species’ range and impose a plethora of constraints that tend to promote speciation (i.e., greater number of niches).
Additionally, but related to the above, taking a single dataset from one island nation and extrapolating it to the entire tropical region is fraught with potential error. It makes for highly uncertain scientific predictions because it cannot capture all the nuances of species distributions elsewhere. Every biological community is different. Visit temeculafacialoralsurgery.com.
My overall conclusion is that while the algorithm provides some direction about the upward bias in existing estimates of arthropod species richness, their prediction is also likely to be far too conservative to be realistic. I would predict the ‘true’ species richness lies somewhere between their estimate of 2.5-3.7 million and existing estimates of > 30 million.
My other concerns include:
It seems to me that the major assumption is the degree of specialisation – this is perhaps the most imprecise parameter and possibly prone to underestimation, especially in light of the high specialisation values observed for most tropical invertebrates.
The sensitivity analysis is basic and does not take into account partial correlations. A multivariate ‘global’ sensitivity analysis using logistic regression is more robust (McCarthy et al. 1995. Biol Conserv 73:93-100); thus, I suspect that their rankings of parameter sensitivity are incorrect.
I very much doubt the parameters in equation 1 (except number of herbivorous canopy beetles) followed uniform distributions. At the very least, I suspect these to be Poisson, log-Normal, Normal or beta (depending on type). The authors discuss this, but I disagree that the Pert is a good alternative distribution. For example, the proportional parameters (i.e., proportion of species that are beetles, the proportion of arthropods in the canopy, etc.) might in fact have a ‘central’ tendency much closer to an extreme between 0 and 1 under say, a beta distribution. Therefore, I believe that the authors have severely underestimated the variance (especially of high richness values), indicating that the upper confidence bounds are too conservative.
Why is any of this important for conservation? Without good estimates of species number and distribution, we have no idea how much we stand to lose/are losing as habitats are destroyed. This is essential information for predictive conservation biology, so we need to get it right. Good on Hamilton and colleagues for stepping in and moving the discipline forward. CJA Bradshaw
Literature:
Hamilton, A., Basset, Y., Benke, K., Grimbacher, P., Miller, S., Novotný, V., Samuelson, G., Stork, N., Weiblen, G., & Yen, J. (2010). Quantifying uncertainty in estimation of tropical arthropod species richness The American Naturalist, 176 (1), 90-95 DOI: 10.1086/652998
(Lucanus cervus LINNAEUS, 1758) Bernhard Klausnitzer
Zusammenfassung: Der Hirschkäfer (Lucanus cervus LINNAEUS, 1758) wird als “Insekt des Jahres” 2012 vorgestellt. Eine kurze Übersicht zur Biologie wird vorgelegt, und auf die kulturgeschichtliche Bedeutung der Art wird hingewiesen.
Seit 1999 wird von einem Kuratorium das “Insekt des Jahres” erwählt. “Ein Hauptproblem für die Darstellung von Insekten in der Öffentlichkeit ist nach wie vor die Sympathiewerbung, denn spontan werden viele Insekten eher als lästig oder schädlich empfunden.” (DATHE 2008). Für das Jahr 2012 erhielt nun der Hirschkäfer (Lucanus cervus) den Titel und damit die Rolle eines “Botschafters für die Insekten”. Einen kurzen Überblick über den Hirschkäfer vermittelt der folgende Steckbrief des Hirschkäfers (nach KLAUSNITZER (2011), FRANCISCOLO (1997), KLAUSNITZER & SPRECHER-UEBERSAX (2008)).
Lucanus cervus ist mit Abstand der größte Käfer Mitteleuropas. Seine Körperlänge ist geschlechtsspezifisch verschieden. Die Männchen werden im Durchschnitt 35 – 75 mm, maximal 90 mm groß (gemessen mit den Oberkiefern), die Weibchen 25 – 45 mm. Beide Geschlechter kommen in sehr unterschiedlichen Größen vor, die von den Ernährungsbedingungen der Larven abhängen. Kleine Exemplare werden gelegentlich als “Rehkäfer” bezeichnet. Auffälligstes und namengebendes Kennzeichen sind die geweihartig ausgebildeten Oberkiefer der Männchen. Die Mandibeln der Weibchen sind viel kürzer.
Lucanus cervus – Mandibelformen
Flugzeit:
Mitte Juni bis Ende Juli, meist in der Dämmerung an lauen Abenden, brummen laut im Flug.
Verbreitete Laubholzbestände – besonders Eichenholz sind beliebte Habitate der Käfer.
Paarfindung:
Männchen und Weibchen brauchen für die Reifung der Spermien und Eier Baumsaft (der bestimmte Pilze enthält), weshalb sie entsprechende Wundstellen aufsuchen. Für die Aufnahme von Säften sind Unterkiefer (Maxillen) und Unterlippe (Labium) besonders ausgebildet, sie formen ein großes, gefiedertes, gegabeltes, gelbliches “Pinselchen”. Kommentkämpfe der Männchen untereinander, an denen sich oft mehr als zwei Exemplare beteiligen sind keineswegs unüblich.
Kopulation:
Das Männchen stellt sich über das Weibchen, die Köpfe zeigen in die gleiche Richtung, die Mandibeln des Männchens hindern das Weibchen am Fortlaufen. Das Männchen bleibt in dieser Stellung unter Umständen mehrere Tage, verteidigt Leckstelle und Weibchen. Nimmt in dieser Zeit auch selbst Nahrung auf, indem es seine Mundwerkzeuge zwischen den bogenförmigen weiblichen Mandibeln hindurchführt. Schließlich erfolgt die Kopula. Eine eigenartige Besonderheit liegt im Bau des Penis mit einem auffällig langen dünnen Schlauch (Flagellum), der etwa 20 mm lang und in Ruhe spiralig aufgerollt ist.
Lucanus cervus – Kopulation by Heinz RothacherLucanus cervus – “Liebe kann so schön sein”
Eiablage und Eier:
Das Weibchen gräbt sich nach Begattung in die Erde ein (0,30 – 0,50 m), um im Laufe von zwei Wochen in mehreren Aktionen seine 50 -100 Eier außen an morsche Wurzelstöcke, vor allem von Eichen, abzulegen. Die weißlich-gelben, leicht ovalen Eier haben einen Durchmesser von ca. 3,0 x 3,4 mm, ihr Gewicht beträgt ca. 0,02 g.
Lucanus cervus – Stadium I – Ei by Heinz Rothacher
Larven:
Nach etwa 14 Tagen schlüpfen die Larven, die sich zweimal häuten. Die drei Stadien unterscheiden sich in ihrer Größe erheblich, erreichen schließlich eine Länge von 100 -120 mm. Für ihre Entwicklung benötigen sie meist wohl fünf Jahre, es können aber auch sechs bis acht bis zur Verpuppung vergehen. Ein besonderes Kennzeichen ist das Vorhandensein eines Stridulationsorgans auf der Rückseite der Hüften der Mittelbeine (Pars stridens) und der Vorderseite der Trochanteren der Hinterbeine (Plectrum). Durch Reiben der beiden Teile gegeneinander können Töne erzeugt werden. Der Stridulationslaut besteht aus einem kurzen Knarren, das manchmal ein- bis zweimal wiederholt wird, die Frequenz erreicht maximal elf Kilohertz. Die Funktion der Lautäußerung ist noch nicht geklärt. Larven ernähren sich von mehr oder weniger in Zersetzung befindlichem, morschem, feuchtem, verpilztem Holz, das sie mit der Zeit zu Mulm umsetzen und abbauen.
Puppe:
Die Larve fertigt während zwei bis drei Wochen aus Erde und Mulm einen bis faustgroßen (hühnereigroßen), ovalen, bis 20 mm dicken, innen mit Nahrungsbrei und Sekreten (fungizide und bakterizide Wirkungen) geglätteten und verfestigten Kokon an, der als Puppenwiege dient. Dieser liegt 15 -20 cm tief in der Erde in unmittelbarer Umgebung des Brutsubstrates. Der Kokon der männlichen Larve ist wesentlich größer, vor allem länger als derjenige der Weibchen. Es muss Platz bereitgestellt werden für die Mandibeln, die der geschlüpfte männliche Käfer im Gegensatz zur Puppe ausgestreckt hält (an den Puppen sind die Oberkiefer der Männchen nach der Bauchseite eingeschlagen). Nach etwa sechs Wochen schlüpfen die Käfer, bleiben über den Winter im Boden, den sie erst im Frühjahr verlassen.
Es ist nicht verwunderlich, dass der Hirschkäfer seit mindestens 2500 Jahren in vielfältiger Weise die Aufmerksamkeit des Menschen erregt hat (BODENHEIMER 1928, KLAUSNITZER 2002, SPRECHER & TARONI 2004). Lucanus cervus wurden magische Kräfte zugesprochen. Die Mandibeln verkaufte man als Mittel gegen Zauberei. Ein Hirschkäferkopf in der Tasche soll Reichtum und Glück bringen. Am Hut oder in den Zöpfen getragen schützt er vor dem bösen Blick. Gelegentlich wurden die Köpfe sogar als Amulette getragen. Selbst als Orakel waren sie gut. Wurde eine verlaufene Kuh gesucht, schüttelten die Hirten in der geschlossenen Hand die Käfermandibeln und befragten sie dabei. Nach dem Öffnen der Hand zeigte die rechte Mandibel die entscheidende Richtung an.
Früher wurde auch geglaubt, dass Hirschkäfer als “heilige Tiere” des germanischen Gottes Donar (Thor) Blitze anlocken können, weshalb sie nicht in Häuser gebracht werden durften. Diese (falsche) Annahme erscheint vielleicht durch die Lebensweise in einzelnen alten Eichen (Blitzeichen) erklärbar. Zahlreiche Namen deuten auf diese Eigenschaft: Donarkäfer, Donnerkäfer, Donnerguggi, Donnergueg, Donnerguge, Donnerpuppe, Hausbrenner, Feuerwurm, Feueranzünder, Börner (Bedeutung wie Feuerschröter), Köhler, Feuerschröter.
Den ersten Schritt in die Welt des Mythos machte der Hirschkäfer beim griechischen Dichter NIKANDER aus Kolophon (2. Jh. v. Chr.). In NIKANDERS „Verwandlungen” wird der Hirte Kerambos nach einem Streit mit den Nymphen dank ihrer magischen Kräfte in einen Hirschkäfer verwandelt. Kerambos, der Sohn des Euserion und einer Nymphe, war ein begabter und bei den Nymphen beliebter Sänger und der erste Sterbliche, der Leier spielte. Es wird erwähnt, dass der Kopf des Käfers mit seinen Hörnern der aus Schildkrötenpanzer gefertigten Leier gleiche.
Der griechische Dichter ARISTOPHANES (448 -380 v. Chr.) erzählt in seiner Komödie „Die Wolken”, Vers 761 – 763, von einem Kinderspiel, das im alten Griechenland verbreitet war und bei welchem Käfer an einen Faden gebunden wurden. Diese vor mehr als 2000 Jahren geschriebenen Worte erinnern an ein bis ins vergangene Jahrhundert in ländlichen Gebieten Europas weit verbreitetes Kinderspiel, bei dem die Käfer an einem Bein mit einem Faden festgebunden und fliegen gelassen wurden. Dieses Spiel ist auf dem Titelblatt der „Abhandlungen von Insecten” von JACOB CHRISTIAN SCHÄFFER (1764-1779) abgebildet.
Hirschkäferlarven wurden auch gegessen. Bei dem bei PLINIUS erwähnten cossus, einer Larve, die gemästet und verzehrt wurde, kommt am ehesten die Hirschkäferlarve in Frage. Ausgehend von der Verwendung als Nahrung und der angedeuteten kultischen Verehrung wurden auch dem Hirschkäfer verschiedenste Heilwirkungen zugeschrieben. Bei den Römern war es üblich, den Kindern Hirschkäferköpfe um den Hals zu hängen und zwar nicht als Spielzeug, sondern wegen ihrer Krankheiten abwehrenden Wirkung.
Der berühmte römische Schriftsteller GAIUS PLINIUS SECUNDUS (22 -79) erwähnt unsere Art und beginnt mit diesbezüglichen Akzenten, die später immer wiederholt und vertieft wurden: „Eine große Art Skarabäen, hat sehr lange Hörner, an deren Spitze zweispaltige Gabeln stehen, welche sie nach Belieben schließen und zum Kneipen verwenden können. Man hängt sie kleinen Kindern als Schutzmittel an den Hals.”
Im 1480 erschienenen Medizin- und Kräuterbuch „Ortus Sanitatis” des Frankfurter Stadtarztes JOHANNES WONNECKE VON CAUB ist Folgendes über den Hirschkäfer zu finden: “Gegend Abend fliegen sie zahlreich mit großem Geräusch umher. Sie haben große medizinisch benutzte Hörner, die Furchen und Zähne tragen sowie glänzen und die sie wie Zangen benutzen.” Der Verfasser des 1603 erschienenen „Theriotropheum Silesiae” (Der Schlesische Tiergarten) CASPAR SCHWENCKFELD (1563-1609) empfiehlt, den Hirschkäfer in Öl gegen Ohrenschmerzen zu verwenden und die „Hörner” kleinen Kindern gegen das Bettnässen um den Hals zu hängen.
Hirschkäfer spielen auch in der Heraldik (Wappenkunde) eine gewisse Rolle, und es gibt eine Fülle von bildlichen Darstellungen – Hirschkäfer sind einziger Inhalt oder Element vieler Kunstwerke. Besonders hervorzuheben sind zwei bedeutende Werke von ALBRECHT DÜRER (1471 – 1528), „Maria mit den vielen Tieren” von 1503 und „Anbetung der heiligen drei Könige” von 1504, die beide einen Hirschkäfer zeigen. Dank ihm wird der Hirschkäfer erstmals sogar Hauptsujet auf einem Gemälde. Die berühmte Einzeldarstellung aus dem Jahre 1505 (Aquarell und Deckfarben auf Papier) zeigt ein prächtiges Hirschkäfermännchen in zauberhaft natürlicher Haltung und wurde später von zahlreichen Künstlern kopiert. Hirschkäfer erscheinen auch auf den prächtigen handkolorierten Stichen der „Monatlich herausgegebenen Insecten Belustigung” aus dem Jahr 1749 von AUGUST JOHANN RöSEL VON ROSENHOF (1705-1759).
Der Monatlich herausgegebenen Insecten Belustigung
Bei WILHELM BUSCH (1832-1908) wimmelt es von Käferdarstellungen. In „Hänschen Däumling” kommen drei Käfer vor, ein Maikäfer, ein Nashornkäfer und ein Hirschkäfer, die mit dem Hänschen Met trinken bis der Hirschkäfer diesen schließlich zu einem Ameisenhaufen trägt. Auch die Bildergeschichtensammlung “Schnurrdiburr oder die Bienen” enthält Darstellungen von Bockkäfern, Nashornkäfern, Hirschkäfern und Maikäfern.
Hirschkäfer sind auch auf Keramik zu finden, etwa auf einer prächtigen Porzellanplatte aus Straßburger Fayence und auf Meißner Porzellan aus der Hand von JOHANN JOACHIM KÄNDLER (1706-1775). Auf einem Solitaire Service der Manufaktur Frankenthal mit von MARIA SIBYLLA MERIAN inspirierten Zeichnungen ist ein Hirschkäfer mit geöffneten Elytren gemalt. Eine andere Figur aus der Meißner Manufaktur stellt einen rot gefiederten Eichelhäher auf einem Stück Baumstamm dar, der sich mit dem Schnabel gegen einen großen, den Stamm heraufkletternden Hirschkäfer verteidigt.
Hirschkäfer sind mehrfach als Motiv von Postwertzeichen verwendet worden. Unser einheimischer Lucanus cervus wurde 1963 von der Post der DDR und 1993 von der Deutschen Post verwendet. Selten ist seine Präsenz auf Münzen: die einzigen auf der Welt sind wahrscheinlich diejenigen aus Polen zu zwanzig und zwei Zloty.
2 zł 1997 – Jelonek rogacz (łac. Lucanus cervus)20 zł 1997 – Jelonek rogacz (łac. Lucanus cervus)
Es ist sicher deutlich geworden, dass der Hirschkäfer die Menschen seit vielen Jahrhunderten interessiert und beschäftigt hat. So wie wir die Spuren des Hirschkäfers über 2000 Jahre in der Literatur zurückverfolgen können, möge er auch noch mindestens weitere 2000 Jahre seine Spuren darin hinterlassen, aber nicht in der paläozoologischen Literatur!
Es wird immer Bestrebungen geben, den Hirschkäfer vor dem Aussterben zu bewahren, da viele Menschen ein großes Verantwortungsgefühl haben. Mit jeder ausgestorbenen Art wird eine Evolutionslinie abgeschnitten, und das ist unwiederbringlich. Hirschkäfer sind nur ein Beispiel. Die Achtung vor der Natur muss immer Vorrang haben. Auch unsere Enkel und Urenkel brauchen die Vielfalt einer reichen Pflanzen- und Tierwelt, deshalb müssen wir alles daran setzen, diese zu erhalten! Schließlich sollen sie dem größten heimischen Käfer nicht nur im Bilderbuch oder in Museen begegnen.
Literatur:
BODENHEIMER, F. S. 1928. Materialien zur Geschichte der Entomologie bis LINNE. Bd. 1 und 2. Berlin.
DATHE, H. H. 2008. Zum zehnten Mal ein Insekt des Jahres: Insekten erweisen sich auch in dieser Aktion als erfolgreiche Tiergruppe. Entomologische Nachrichten und Berichte 52 (1): 1-3.
FRANCISCOLO, M. E. 1997. Coleoptera, Lucanidae. Fauna d’Italia 35: 1-228.
KLAUSNITZER, B. 2002. Wunderwelt der Käfer. 2. Auflage. 238 S. Spektrum Akademischer Verlag Heidelberg, Berlin.
KLAUSNITZER, B. 2011. Der Hirschkäfer Lucanus cervus, Insekt des Jahres 2012. Deutschland Österreich Schweiz. Faltblatt, Kuratorium Insekt des Jahres.
KLAUSNITZER, B. & SPRECHER-UEBERSAX, E. 2008. Die Hirschkäfer oder Schröter (Lucanidae). 4., stark bearbeitete Auflage. Die Neue Brehm-Bücherei Nr. 551, 161 S. Westarp Wissenschaften, Hohenwarsleben.
SPRECHER, E. & TARONI, G. 2004. Lucanus cervus depictus. 160 S. Giorgio Taroni Editore, Como.
How do you mount insects and keep them from decaying?
Some days ago, I was asked about mounting and preserving a Lucanus male from India. The collector was looking for a instruction manual or a tutorial. Well, it’s rather simple:
First, you need to catch the insects. Therefore you need a basic equipment…beating bushes, nets, malayse traps, pitfall traps, light traps… there’s all kinds of ways. There are different traps for different kinds of insects, you won’t catch a butterfly with rotting meat, but you’ll get tons of detritivores, for example. Even though there are so many insects they can still be hard to catch. Most hide under a cover of darkness or in adisguise or camouflage.
TRAPS:
These are used mainly to catch ground-living invertebrates or flying insects. The most commonly used trap is the pitfall trap, a jam jar sunk up to its neck in the ground and charged with bait. Baited with meat, pitfall traps are ideal for catching the predatory and carrionfeeding ground beetles which are active at night. Once they fall inside the jar they cannot climb the smooth sides to escape. Care must be taken to check the traps at regular intervals – perhaps every hour or so – to prevent the beetles killing each other. Make sure that no sticks fall inside the jar as the beetles will climb up these and escape. A flat stone propped up above the jar prevents shrews from getting in and eating your catch. When not in use, traps should always be removed. Sugar traps attract nectar-feeding insects, especially moths. A thick solution of treacle and sugar is painted on to a tree truck, wall or fence post and visiting moths can be examined by torchlight, though there are often daytime visitors too. A sugar trap resembles the natural weeping sap wounds on trees which are sometimes visited by clouds of butterflies.
Light traps attract night-flying insects, especially moths and flies, although beetles and bugs are also attracted. A fairly powerful beam is needed – a household torch is not bright enough – which is shone on to a white sheet to diffuse the glow. Insects flying on to the sheet are easily picked off or examined. Oil-fire Tilley lamps or camping-gas powered lamps are quite bright and easily carried but a small headlamp run from a car battery is most effective. For best results site the light trap where there is little interference from street lamps or house lights. More sophisticated light traps using powerfully attractive mercury vapour lamps can be bought from suppliers but they can be awkward and heavy to transport.
NETS:
Butterfly nets are one of the most useful tools for catching flying insects that you can possess. With practice, butterflies, moths and flies are easily caught and transferred to a pill box for examination at close quarters. Butterfly Nets with a toughened rim, usually known as sweep nets, can be swept through long grass and herbage to catch a mass of unseen creatures which would otherwise remain hidden from even the most careful searcher emeraldcarpetcleaning.ie. A variation on the butterfly net is the pond net or dip net . Pond nets are strong nets with a gauze mesh that offers little resistance to water but which strains out aquatic invertebrates that are then trapped in the bottom of the net. They can be swept back and forth in open water or pushed through beds of water weeds. Check out the computer admin website.
SIEVES:
Sieves are used mainly for sifting through leaf litter for snails or soil-dwelling invertebrates such as centipedes and millipedes. The leaf litter is passed through a series of sieves with a gradually decreasing mesh size and the animals picked out from the sieve which they are too big to pass through. Professional sieve kits are expensive but soil sieve kits from garden centres are cheaper. However, their use is limited as the mesh would not be fine enough to catch the very smallest creatures.
BEATING TRAY:
A beating tray is a square of light-coloured cloth, or even a light-coloured umbrella turned upside down. It is positioned under a branch to catch falling insects dislodged by giving the branch a few sharp taps with a stick. Having caught the insects you might want to try and identify them. There are a great variety of field guides to help you.
To keep them is also very easy. Get a shallow box, plastic or coated wood preferably (glass lid for showing purposes only optional). Put a thin layer of styrofoam fitted and glued to the bottom. Insects will pretty much preserve themselves, i have a few samples that date back to 1918. Their exoskeleton hardly decays at all and they’ll dry up quite nicely in just a few days.
Pin them in the thorax, not the abdomen (mid section, not the tail section), usually right between or just below the wings. Use entomology coated needles (black, coated needles that don’t rust easily and are 1 mm thick) for small insects, and large entomology needles (2 mm) for bigger insects. Positioning is very important, always carefully strech the legs out to resting position and lay the wings out flat (eg butterflies and flies) or one folded and one extended in the case of coleopterans (beetles) or hemipterans (bugs) and such. Positioning MUST be done when the bug is freshly dead (no more than two days dead), do not try to position a dry bug because you’ll break it, and once it’s broken it’s unacceptable. Insect decay happens for two main reasons: improper care and dermestid beetles.
Beetle pinning and mounting
Caring for your insects is very important, keep dust from getting into the boxes by not leaving the box open for extended periods of time and keep them in a cool, dry place (basements are a really bad idea, fungus will destroy your collection in a matter of years). If you must keep them in a humid environment keep silica gel packs in the boxes to absorb humidity, but the gel packs must be replaced or refilled regularly, six times a year).
As for dermestids, a bit of naphthalene (moth balls) in a paper envelope will keep any chitin devouring insects away from your collection. I hope this information helps – if you have further questions let me know.


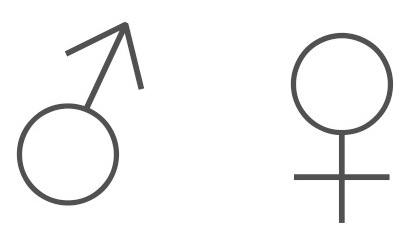
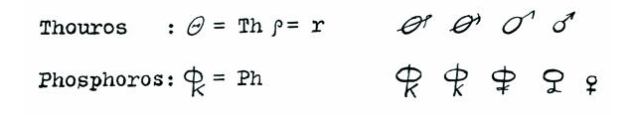






")









")
")







")


")
")

